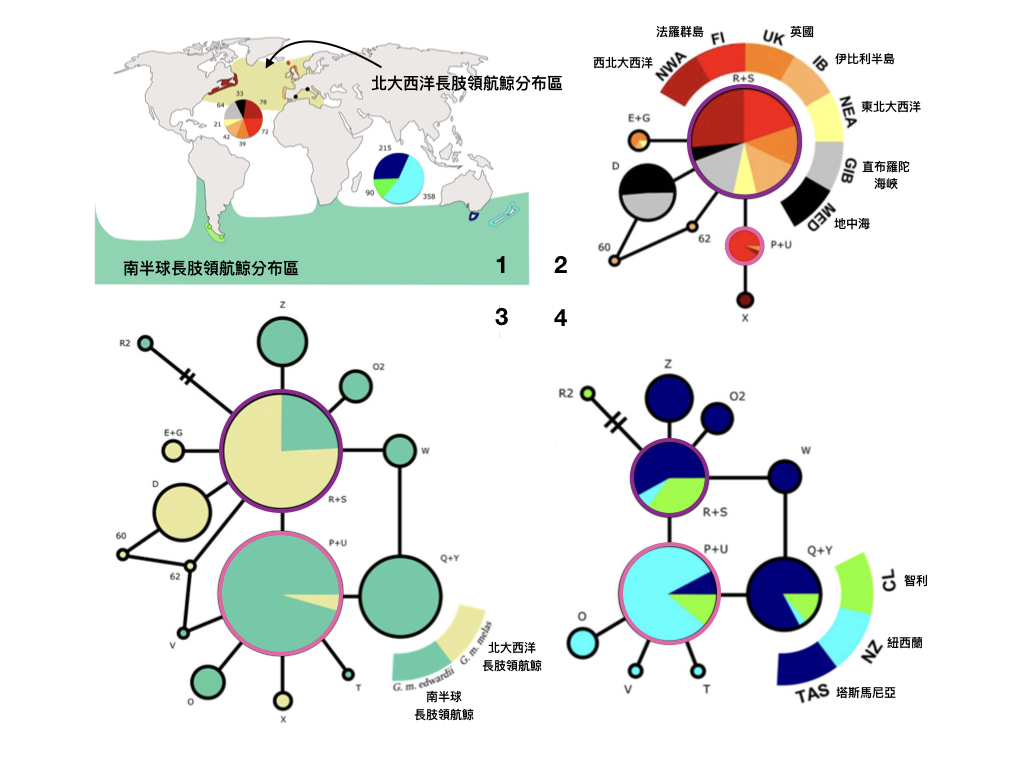
但是啊,就像先前系列文章常說的,這個研究僅使用粒線體 DNA 上一個短片段作為分子標記,先不說粒線體 DNA 的母系遺傳特色無法證實不同族群之間有沒有基因流,僅用短序列片段畫出來的譜系樹預測失準的可能性也比較大。但是考慮到跨海域研究鯨豚的困難性,這項研究的發現還是為長肢領航鯨的族群分化史提供了有用的證據。這項研究發表於知名學術出版集團 Nature 旗下的開放期刊,「Scientific Reports」。
說了這麼多,那世界上的長肢領航鯨是不是還是分成兩個亞種?
結論是「是的」,因為即便南北半球的長肢領航鯨有共有的粒線體 DNA 基因型,這些基因型在兩邊出現的頻率明顯不一樣,顯然兩群長肢領航鯨之間的基因流已經不復存在。一篇去年 (2019 年) 發表在「Marine Mammal Science」的研究也顯示,南北大西洋海域的長肢領航鯨,在頭骨形態測量值上有明顯的差異。再加上赤道熱帶海域這麼明顯的地理屏障,所以在更有力的近期基因交流證據出現之前,這兩個亞種的地位應該不會有所改變。
參考資料:
Kraft, S., Pérez-Álvarez, M., Olavarría, C., & Poulin, E. (2020). Global phylogeography and genetic diversity of the long-finned pilot whale Globicephala melas, with new data from the southeastern Pacific. Scientific Reports, 10(1), 1-13.
Marina, T. I., Marchesi, M. C., & Goodall, R. N. P. (2019). Long‐finned pilot whale (Globicephala melas, Traill 1809) subspecies in the Atlantic Ocean: Are there differences in their skulls? Marine Mammal Science 35(2), 660–676.
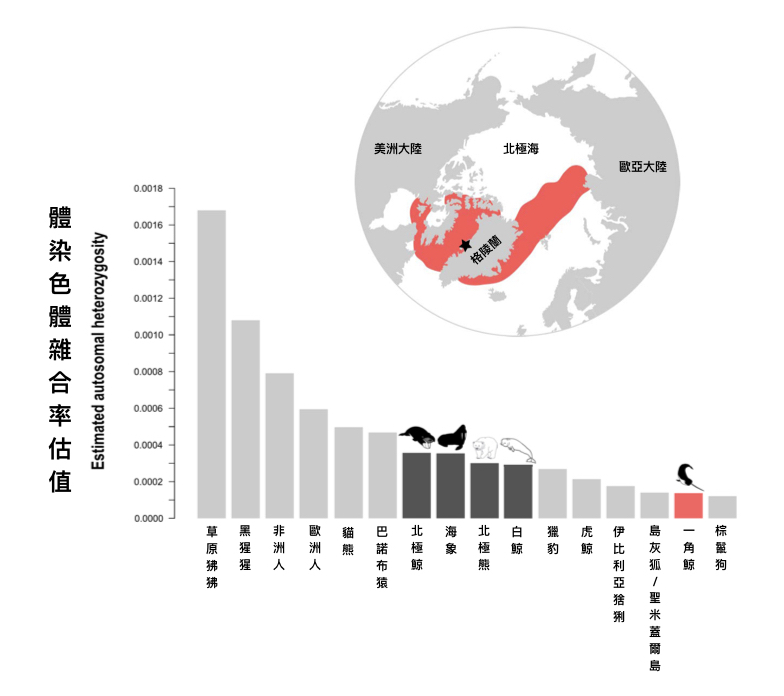
圖二:上圖為一角鯨的分布圖 (紅色範圍),黑色星星為本研究樣本採樣地點。下圖為各種動物的體染色體雜合率估計值比較圖,鐵灰色為一角鯨在北極的快樂夥伴們的體染色體雜合率估計值。體染色體雜合率越高代表基因多樣性越高、有效族群量 (effective population size) 越大。圖片來源:Figure 1 in Westbury et al. (2019) iScience, 15, 592-599.
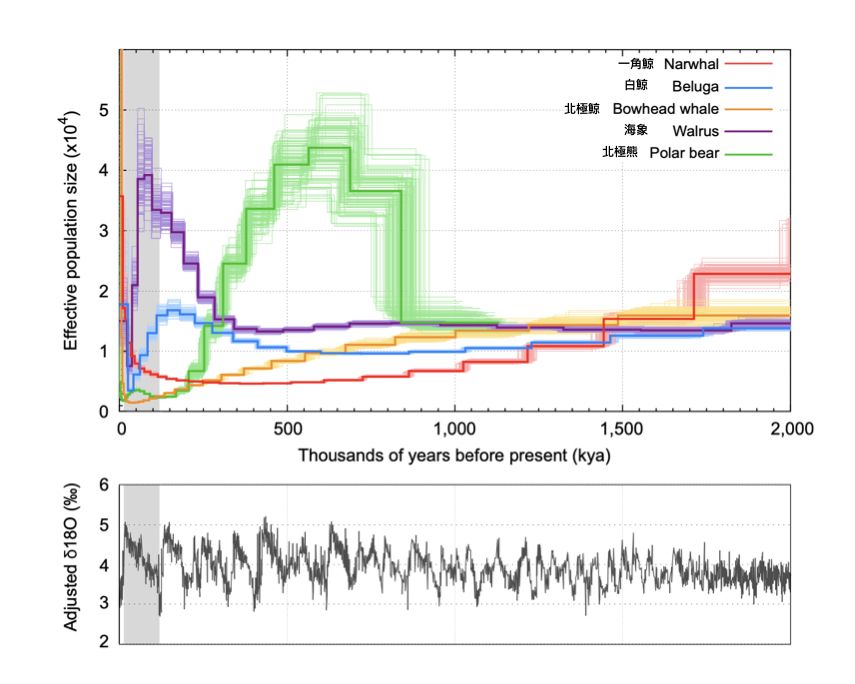
圖三:利用基因體資料推算出來的各種北極動物的有效族群量 (effective population size) 歷史動態變化 (上圖),以及對照的歷史氣候變化 (下圖)。橫軸單位均為距今千年前 (kya)。上圖的有效族群量代表該世代中成功生殖下一代的個體數目 (而非實際族群數量 — 因為並不是每一隻個體都生得出小孩),而下圖縱軸的單位 delta-18O 表示各個時期氧穩定同位素的比例;數值越高表示環境溫度越低,數值越低表示環境溫度越高。圖中灰色區塊表示最近一次冰河期的時間 (距今 11—2 萬年前)。圖片來源:Figure 4 in Westbury et al. (2019) iScience, 15, 592-599.
Lowry, L., Laidre, K. & Reeves, R. 2017. Monodon monoceros. The IUCN Red List of Threatened Species 2017: e.T13704A50367651. Downloaded on 25 October 2019.
Westbury, M. V., Petersen, B., Garde, E., Heide-Jørgensen, M. P., & Lorenzen, E. D. (2019). Narwhal Genome Reveals Long-Term Low Genetic Diversity despite Current Large Abundance Size. iScience, 15, 592-599.
流冰虎鯨 (又稱大 B 型虎鯨,Large Type B; Pack Ice killer whale)
和 A 型虎鯨差不多大小,體色主要以灰白雙色組成,在光線夠的時候,可以觀察到深灰色的披肩 (dorsal cape)。因為被海藻附身的關係,很多個體會看起來黃黃髒髒的。眼斑很大、角度與體軸平行。主要出沒在環南極有冰的海域,因為他們主要以棲息在冰塊上的海豹們為主食。這個類型的虎鯨也會在南半球高低緯度海域間進行快速遷徙,不過大部分的時間都待在南極海域。
傑朗奇虎鯨 (又稱小 B 型虎鯨,Small Type B; Gerlanche killer whale)
這個類型的虎鯨體型較流冰虎鯨 (大 B 型) 小 1 公尺、體重輕一半。體色外型跟流冰虎鯨幾乎一樣:有深灰色披肩、大又平行體軸的眼班、黃黃的海藻裝束。傑朗奇虎鯨 (小 B 型) 主要以魚類、頭足類為食,有時候也吃企鵝,主要在無冰的海域出沒。他們也會在南半球高低緯度海域間進行快速遷徙,不過同樣大部分的時間都待在南極海域。
羅斯海虎鯨 (又稱 C 型虎鯨,Type C; Ross Sea killer whale)
體型比北半球虎鯨小大概 1–3 公尺左右的侏儒型虎鯨。體色外型跟流冰虎鯨、傑朗奇虎鯨等 B 型虎鯨相似,一樣有深灰色披肩、黃黄藻類附身的特色,但是眼斑比較狹長,和體軸呈 45° 角向前傾斜,乍看之下像是隨時都在生氣的樣子。目前已知只會出現在南極大陸東側,以南極鱈魚 (Dissostichus mawasoni) 等魚類為食。和 B 型虎鯨也會快速遷徙到低緯海域,但是整年大部分的時間主要都在南極附近活動。
最新的分子遺傳分析結果大致上支持這樣的分群系統;兩種 B 型虎鯨、C 型虎鯨和 D 型虎鯨,各自形成獨立的演化支系 (詳情請見上一集的圖一),而且彼此間已經有很長一段時間沒有基因交流。有些學者甚至主張 C 型和 D 型虎鯨和其他虎鯨分家的時間已經長達 15 萬年,應該足以視為獨立的物種。然而 A 型虎鯨的結果就不像其他類型那樣「整齊」,目前的分子遺傳研究顯示 A 型虎鯨,可能是來自於多個演化支系的後代組合。
但是啊,北大西洋這兩型虎鯨之間的遺傳差異,並不像太平洋或南極的諸多虎鯨類群那樣壁壘分明。近年來研究完整粒線體基因體序列的研究發現,第二型虎鯨可能和南極的 A 型虎鯨一樣,是由多個演化支系的虎鯨綜合而成;而第一型虎鯨裡,似乎還可以再細分出偏好鯡魚和偏好鮪魚等不同的家族支系。所以說虎鯨到底有幾種?或許要等更多來自於西太平洋、南太平洋、南大西洋以及印度洋等等海域的調查資料出爐後,才能真正下定論吧。
參考資料
Durban, J. W., Fearnbach, H., Burrows, D. G., Ylitalo, G. M., & Pitman, R. L. (2017). Morphological and ecological evidence for two sympatric forms of Type B killer whale around the Antarctic Peninsula. Polar Biology, 40(1), 231-236.
Foote, A. D., Newton, J., Piertney, S. B., Willerslev, E., & Gilbert, M. T. P. (2009). Ecological, morphological and genetic divergence of sympatric North Atlantic killer whale populations. Molecular Ecology, 18(24), 5207-5217.
Foote, A. D., Vilstrup, J. T., De Stephanis, R., Verborgh, P., Abel Nielsen, S. C., Deaville, R., … & Piertney, S. B. (2011). Genetic differentiation among North Atlantic killer whale populations. Molecular Ecology, 20(3), 629-641
Foote, A. D., Vijay, N., Ávila-Arcos, M. C., Baird, R. W., Durban, J. W., Fumagalli, M., … & Robertson, K. M. (2016). Genome-culture coevolution promotes rapid divergence of killer whale ecotypes. Nature Communications, 7, 11693.
Hoelzel, A. R., Natoli, A., Dahlheim, M. E., Olavarria, C., Baird, R. W., & Black, N. A. (2002). Low worldwide genetic diversity in the killer whale (Orcinus orca): implications for demographic history. Proceedings of the Royal Society of London. Series B: Biological Sciences, 269(1499), 1467-1473.
Jefferson, T. A., Webber, M, & Pitman, R. (2015). Marine Mammals of the World. 2nd Edition. Academic Press. 616 Pp.
Morin, P. A., Archer, F. I., Foote, A. D., Vilstrup, J., Allen, E. E., Wade, P., … & Bouffard, P. (2010). Complete mitochondrial genome phylogeographic analysis of killer whales (Orcinus orca) indicates multiple species. Genome research, 20(7), 908-916.
Pitman, R. L., and P. Ensor. 2003. Three forms of killer whales (Orcinus orca) in Antarctic waters. Journal of Cetacean Research and Management 5:131–140.
Pitman, R. L., J. W. Durban, M. Greenfelder, et al. 2011. Observations of a distinctive morphotype of killer whale (Orcinus orca), type D, from subantarctic waters. Polar Biology 34:303–306.
BBC News. (2018). Moray Firth bottlenose dolphin numbers ‘stable’ says SNH. https://www.bbc.com/news/uk-scotland-north-east-orkney-shetland-43554134 (accessed 2 April 2019)
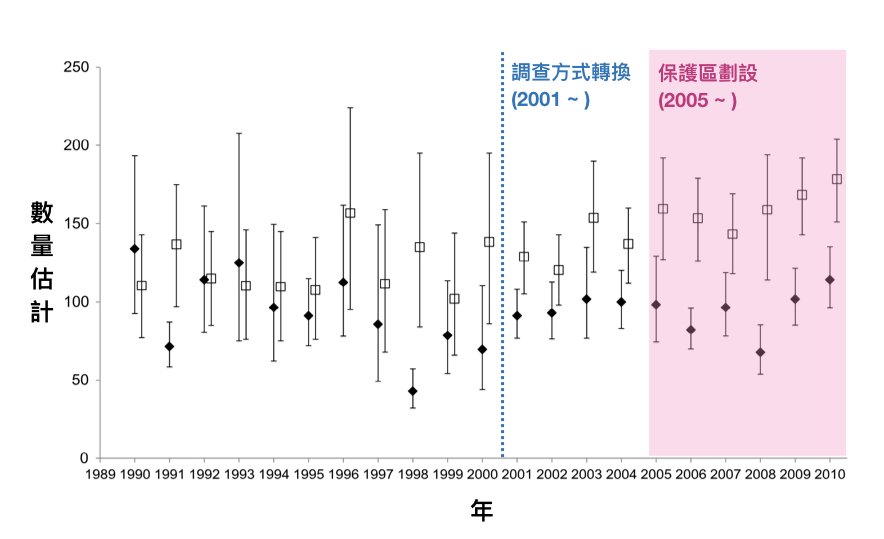
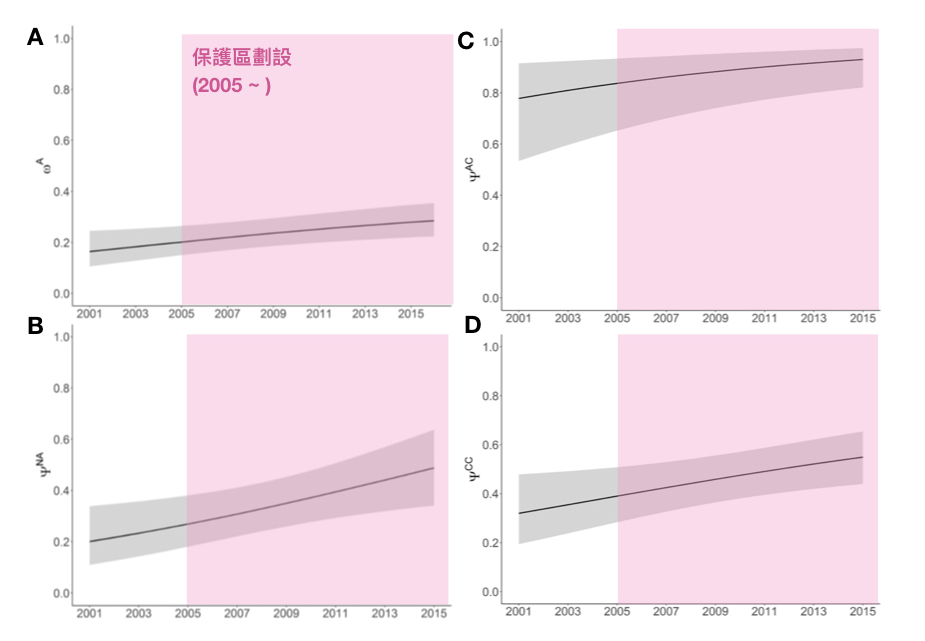
Cheney, B., Corkrey, R., Durban, J. W., Grellier, K., Hammond, P. S., Islas-Villanueva, V., … & Wilson, B. (2014). Long-term trends in the use of a protected area by small cetaceans in relation to changes in population status. Global Ecology and Conservation, 2, 118-128.
Cheney, B. J., Thompson, P. M., & Cordes, L. S. (2019). Increasing trends in fecundity and calf survival of bottlenose dolphins in a marine protected area. Scientific Reports, 9(1), 1767.
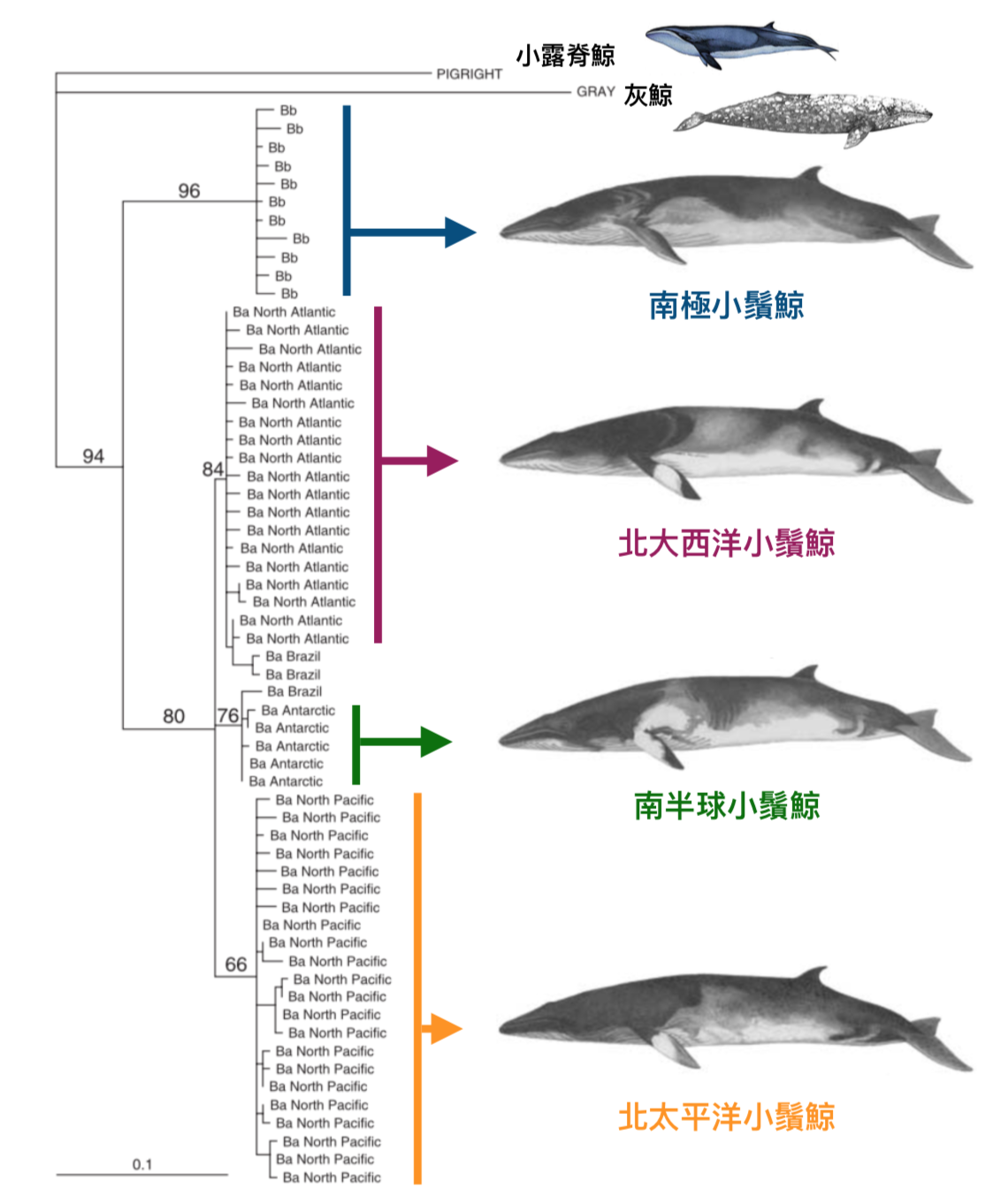
圖一:利用 340bp 粒線體 DNA control region 序列多樣性重建的小鬚鯨親緣譜系圖。該譜系樹以小露脊鯨與灰鯨為外群,分支上的數字表示該支系成立的可信度 (quartet puzzling reliability values, 最大值為 100)。請注意圖中鯨魚大小並未以真實比例呈現。圖片來源:Figs. 1 & 4 in Pastane et al. 2007. Mol. Ecol. 16: 1481–1495.
Pastene, L. A., Goto, M., Kanda, N., Zerbini, A. N., Kerem, D. A. N., Watanabe, K., … & Palsböll, P. J. (2007). Radiation and speciation of pelagic organisms during periods of global warming: the case of the common minke whale, Balaenoptera acutorostrata. Molecular Ecology, 16(7), 1481-1495.
Pastene, L. A., Acevedo, J., Goto, M., Zerbini, A. N., Acuna, P., & Aguayo-Lobo, A. (2010). Population structure and possible migratory links of common minke whales, Balaenoptera acutorostrata, in the Southern Hemisphere. Conservation Genetics, 11(4), 1553-1558.
Perrin, W. F., Mallette, S. D., Brownell JR., R. L. (2018) Minke whales. In: Würsig, B., Thewissen, J. G. M., Kovacs, K. M. (eds.) Encyclopedia of marine mammals. Third Edition. Academic Press.