(原文刊載於台灣大學科學教育發展中心 CASE 報科學部落格,2018 年 12 月 6 日)

撰文|陳瑩
不知道各位讀者們是否還記得,筆者在先前的文章<鬚鯨的吃相> 中提過、那些沒有牙齒的海洋巨獸「鬚鯨」?
在美國南北戰爭打得如火如荼的 1864 年,時任大英博物館動物學部主任的英國皇家院士格雷 (John Edward Gray, 1800–1875),收到了一組遠從南台灣寄來的鬚鯨骨骼標本,請求鑑定物種。寄件人是當時的英國駐台代表,同樣是知名生物學家的 R. 斯文豪 (Robert Swinhoe, 1836—1877)。
這具擱淺鬚鯨的標本,只包括了部分頭骨、三個頸椎、第一節和其他七個位置不明的脊椎骨,以及八根肋骨 (圖一)。由於這些部分的骨骼在諸多鬚鯨物種之間極為相似,所以就算是鯨豚大師格雷,做這份鑑定工作仍然是困難重重。
隔年,格雷將這具標本命名為斯氏鯨 (Balaenoptera swinhoei)。他認為這具標本應該是鬚鯨屬 (Balaenoptera) 的物種,雖然說有諸多特徵和長須鯨 (Balaenoptera physalus) [註] 非常相似。如果標本齊全的話,應該就能確認其特殊性。1868 年,格雷甚至把斯氏鯨「升級」成獨立一屬的物種 Swinhoia chinesis,就像他在 1846 年時,將大翅鯨從鬚鯨屬中「升級」成為大翅鯨屬 (Megaptera)裡單一物種,Megaptera novaeangliae 一樣。
然而,在一百五十多年後的今年 (2018 年),一篇發表在海洋哺乳動物學會的期刊「Marine Mammal Science」的論文,以古 DNA 鑑定技術,證實這具現存於英國自然史博物館的斯氏鯨模式標本,其實是一隻大翅鯨,而且是一隻屬於北太平洋最常見的大翅鯨支系,「庫吉拉大翅鯨 (M. n. kuzira)」的鯨魚。
格雷的誤判其實情有可原。除了標本本身不完整以外,最新的研究結果顯示,大翅鯨、長須鯨和其他鬚鯨,在基因結構上頗為相似,而且在過去數百萬年種化過程中,有過一段無法忽視的雜交歷史。
一組由瑞典和德國科學家組成的研究團隊,利用次世代基因定序技術,針對長須鯨、藍鯨、鰮鯨 (亦稱「塞鯨」)、小鬚鯨、大翅鯨和灰鯨等六種鬚鯨的基因體 (genome) 多樣性進行演化親緣分析。過去傳統分子遺傳學的定序方法,僅能對粒線體或細胞核內的特定 DNA 片段進行多樣性解析,在缺乏大量鬚鯨樣本的情況下,分析結果經常像瞎子摸象一樣,無法完整呈現物種的親緣圖譜。現在有了全基因體定序技術,即使每個種類的鯨魚數量不多,透過大規模比對不同鯨種的基因相似性,也已經足夠解決物種之間演化歷史這個層級的科學問題。
透過分析超過三萬四千個基因體片段,研究人員結果發現,在傳統形態學分類中,總是和藍鯨、鰮鯨、小鬚鯨歸為同屬的長須鯨,其實和被歸類為獨立屬的大翅鯨和灰鯨,有著比較相似的基因圖譜。此外,小鬚鯨的基因也沒有和藍鯨、鰮鯨等「同屬物種」比較相似;其中的基因差異,甚至大於大翅鯨與藍鯨「異屬物種」之間的差別 (圖二)。也就是說,現行根據解剖形態學特徵的分類方法,似乎無法反映鬚鯨的演化歷史。如果以基因體的資料為依歸,那麼除了小鬚鯨之外,其他的鬚鯨應該大致可以歸為「藍鯨/鰮鯨」、「長須鯨/大翅鯨」以及「灰鯨」三大類群。
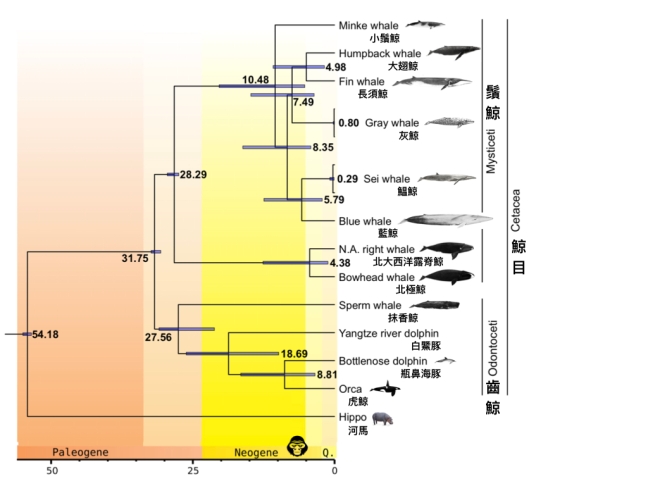
同樣是使用基因體資料,研究人員推估,鬚鯨們大概是在距今 10.5—7.5 百萬年前開始快速輻射種化,原因也許和當時地球洋流改變有關。不過,在鬚鯨種化的過程中 (直到今日),一直有著異種混血的現象。特別是藍鯨——堪稱是鬚鯨界的混血天王/天后——幾乎和其他五個物種都有過一段雜交的歷史。這樣說來,日前冰島捕鯨人抓到一隻藍鯨和長須鯨混血鯨魚、在鯨豚保育圈裡鬧得沸沸揚揚的事件,似乎也沒有那麼奇怪了。
研究人員認為,不同演化支系的鯨魚能夠在頻繁雜交中各自演化成自成一格的物種,顯示同域演化 (sympatric evolution) 這個種化模式,也許在缺乏明顯地理屏障的海洋環境中頗為重要。
這項研究發表於今年四月出刊的開放期刊「Science Adavances」。
可惜的是,這項研究中並沒有包括一些主要出現在印度-太平洋地區的鬚鯨,像是布氏鯨、大村鯨 (角島鯨) 等等種類。雖然說先前的形態學和早期的分子遺傳研究顯示,布氏鯨和大村鯨可能與鰮鯨有著親近的血緣關係,因此極有可能也和藍鯨是屬於同一個類群。但是是否真是如此,大概還是需要更精緻的分析資料支持。
註:當時的長須鯨是獨立於長須鯨屬 (Physalus) 之下的物種,今日則和其他多數的鬚鯨一樣,屬於鬚鯨屬 (Balaenoptera) 的物種。是說長須鯨屬這個分類群,其實也是格雷創立的。
參考資料:
- Árnason, Ú., Lammers, F., Kumar, V., Nilsson, M. A., & Janke, A. (2018). Whole-genome sequencing of the blue whale and other rorquals finds signatures for introgressive gene flow. Science Advances, 4(4), eaap9873.
- Archer, F., Robertson, K., Sabin, R., & Brownell, R. L. (2018). Taxonomic status of a “finner whale”(Balaenoptera swinhoei Gray, 1865) from southern Taiwan. Marine Mammal Science. https://doi.org/10.1111/mms.12496
- Gray, J. E. (1865). XVII.—Notice of a new Finner Whale from Formosa. Annals and Magazine of Natural History, 16:93, 148-148, DOI: 10.1080/00222936508679398
- Gray, J. E. (1866). Catalogue of seals and whales in the British Museum (Vol. 1). Order of the Trustees.
- McGrath, M. (2018). Whale killing: DNA shows Iceland whale was rare hybrid. BBC News, 20 July 2018. https://www.bbc.com/news/science-environment-44809115. <accessed on 12 October 2018>
- Sasaki, T., Nikaido, M., Wada, S., Yamada, T. K., Cao, Y., Hasegawa, M., & Okada, N. (2006). Balaenoptera omurai is a newly discovered baleen whale that represents an ancient evolutionary lineage. Molecular Phylogenetics and Evolution, 41(1), 40-52.